
Introduction
Pancreatic ductal adenocarcinoma (PDAC) occupies a uniquely hostile position in the oncological landscape, responsible for a less than 10% 5 year survival rate and near recurrence in just 14 months in more than 80% patients. Characterized by a dense, desmoplastic stroma, chemo-resilience, and profound immune exclusion “cold tumor morphology”, PDAC has long resisted the therapeutic advances that have transformed outcomes in other malignancies. Its designation as an immunologically ‘cold’ tumor reflects not merely a quantitative deficit of infiltrating lymphocytes because of a smaller number of passenger mutations rendering less opportunity for immune cells to recognize, but a qualitative failure of immune recognition, a microenvironment engineered, through evolutionary selection pressure, to neutralize the host’s most potent cellular defenses. Against this formidable backdrop, cancer vaccines represent an ingenuity that escapes the traditional architecture of macabre cytotoxic therapies and palliative care and brings into conversation something more conceptually ambitious: the active programming of immune memory against cancer which unlike targeting the tumor directly, seek to train the adaptive immune system to imprint specificity on T cells of those antigens lurking in the body, so that immunity becomes, in effect, a living therapeutic.
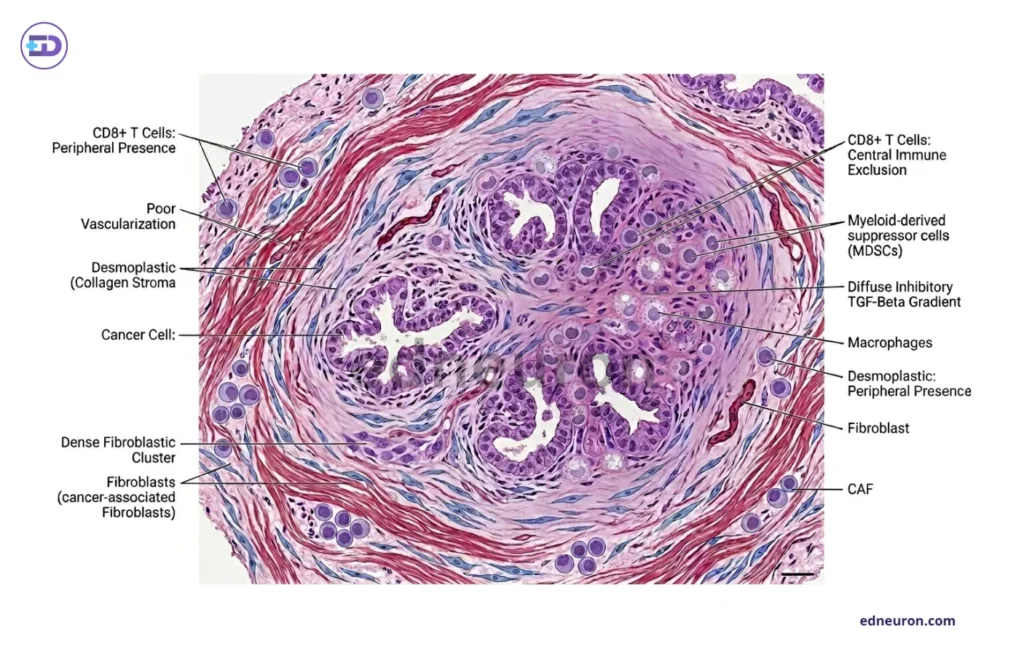
Fig 1: Histopathological diagram of a pancreatic cold tumor-notice the dense collagenous stroma and immunosuppressive microenvironment.
Types of Pancreatic Cancer Vaccines
Contemporary vaccine strategies for PDAC span a spectrum from highly individualized to broadly applicable ones. Personalized mRNA vaccines, most notably autogene cevumeran, are synthesized from more than 20 major histocompatibility complex class 1- and 2-restricted tumor-specific neoantigen sequences derived from genomic profiling of each patient’s resected tumor. This patient-specific, tailored mRNA vaccine represents the highest degree of immunological precision currently achievable. Dendritic cell (DC) vaccines, on the other hand, use patients’ own DCs that are primed and pulsed ex vivo with tumor-associated antigens. These are then reinfused into sentinel lymph nodes, where DCs (important APCs) process and present antigens via MHC molecules, thereby triggering an activation cascade of CD8+ and CD4+ cells to facilitate tumor destruction. Whole-cell vaccines- GVAX, employ allogeneic whole tumor cells engineered to secrete granulocyte-macrophage colony-stimulating factor (GM-CSF), generating a broad antigenic stimulus capable of significantly populating the tumor with diverse cytotoxic T-cells. This neoadjuvant GVAX in combination with chemoradiotherapy has proven better relapse-free survival and overall survival in these PDAC patients. Each platform embodies a different balance between specificity, cost, and manufacturing feasibility, a tension that will define their clinical trajectories.
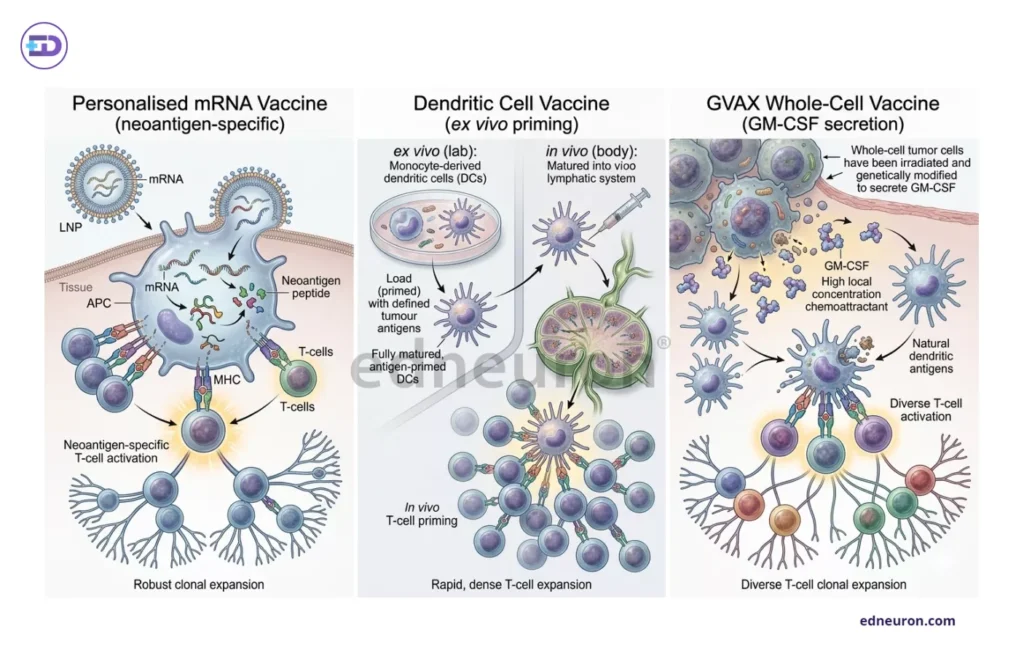
Fig 2 : Different types of vaccines lead to subtle differences in types of T cell activation it produces.
Mechanism of These Vaccines
The problem with PDAC is low passenger mutation rates and an immunosuppressive microenvironment that retards immune cell recognition and infiltration, allowing these malignant cells to proliferate in impunity, veiled under an oblivious immune system. The core logic underlying these vaccines does not stem from something foreign but rather reflects a common mechanism we all know. mRNA vaccines are produced from various neoantigens, which are combined with MHC-restricted molecules. MHC class I molecules process and present endogenous antigens. This activates CD8+ cytotoxic T lymphocytes, which can directly eliminate tumor cells. Meanwhile, MHC class II engagement recruits exogenous antigen-presenting CD4+ helper T cells, which boost the immune response and sustain the humoral response. Specifically, in PDAC, this process is impeded at multiple nodes by transforming growth factor-beta (TGF-β)-mediated stromal immunosuppression, the accumulation of myeloid-derived suppressor cells (MDSCs) within the tumor microenvironment driven by KRAS mutations, and by the progressive exhaustion of tumor-infiltrating lymphocytes. A significant recent advance has been the integration of artificial intelligence into neoantigen discovery pipelines. For a cancer like PDAC which exhibits a comparatively low tumour mutational burden, reducing the probabilistic yield of immunogenic neoantigens, the traditional 6-8 weeks to create a vaccine risking losing the therapeutic window, and the extensive heterogeneity leading to scarcity of targetable antigens, AI-driven multi-omics platforms, as described by Zhang J et al., address this constraint by modelling human leukocyte antigen (HLA)-T cell receptor (TCR) binding affinity with greater precision to these neoantigens, identifying cryptic epitopes that conventional algorithms overlook thereby producing polytopic respnoses. Additionally, matrix metalloproteinase-9 (MMP-9)-targeted lipid nanoparticles (LNPs) have been explored to improve vaccine delivery across the stromal barrier, enhancing lymphocyte infiltration.
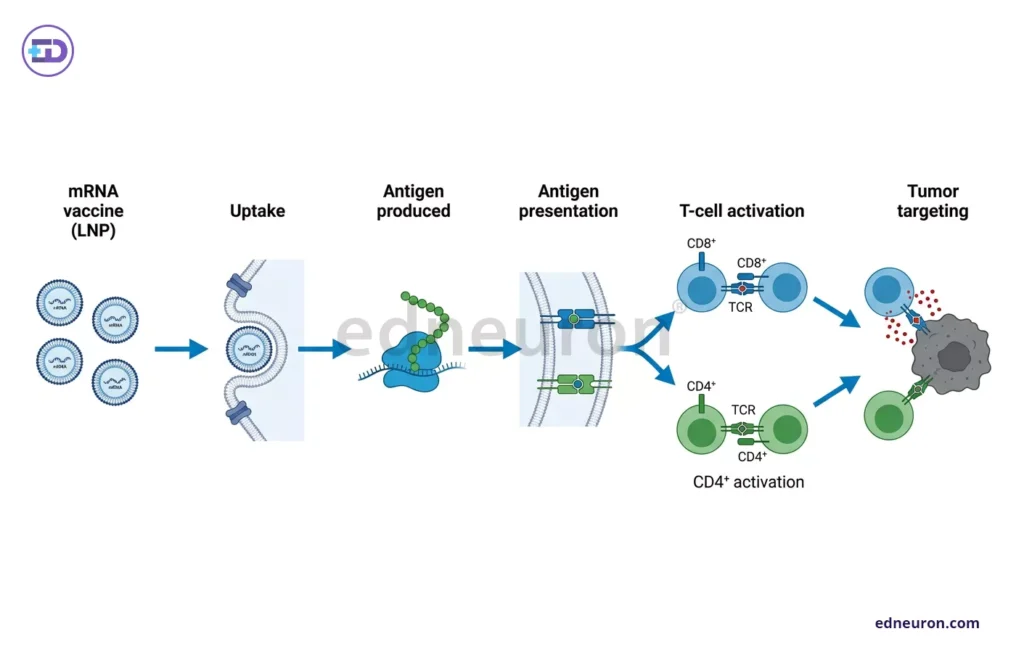
Fig 3 : Simple representation of mechanism of mRNA vaccine activating the T cells to target tumor destruction.
Clinical Evidence and Outcomes
The phase 1 trial of autogene cevumeran by Rojas LA et al. reports such staggering, impactful results- among 16 patients with resected PDAC who received the personalized mRNA vaccine in combination with atezolizumab and mFOLFIRINOX, eight (50%) mounted measurable, high-magnitude neoantigen-specific T-cell responses. Half of these responses were found to be polytopic against at least one vaccine neoantigen. One limitation was perhaps the trials’ modest size, which required interpretive caution. But even then, it does not in any way undermine the strength of these responses or the clear survival difference between responders and non-responders. These findings are noteworthy and provide strong proof of concept, surpassing historical norms for the treatment potential of engineered tumor immunity. Such datasets suggest that the immunological response rate hovering around 50% may serve as a surrogate marker for meaningful clinical benefit. However, the absence of phase 3 validation remains the critical evidentiary gap.
Recent Advances
The period between 2023 and 2026 has witnessed an array of innovations in computational and materials science that are reshaping vaccine design and clinical workflows. AI-powered neoantigen modeling by mining whole-exome sequencing, transcriptomic, and HLA-typing data simultaneously and accurately determining the timing of combined therapy before lymphocytic exhaustion occurs, these platforms can construct vaccine targets with a precision that manual curation cannot replicate. Lipid nanoparticle delivery systems, adapted from the mRNA vaccine infrastructure developed during the COVID-19 pandemic, have enabled more efficient cytosolic antigen delivery and enhanced the priming of long-lived memory CD8+ T cells. With the triumph in the phase 1 trial, the phase 2 expansion of the autogene cevumeran program, confirmed in 2026, marks an important milestone. This signals sufficient phase 1 efficacy to justify broader investigation and, with it, the first rigorous test of whether personalized mRNA vaccination can achieve a generalizable survival benefit.
Challenges and Limitations
Despite advances in immunotherapeutic sciences, PDAC’s reputation as a perfidious entity stems from multidimensional resistance to obliteration that is not easily dismantled. The tumor microenvironment is not just a mere physical barrier but an active immunosuppressive organ secreting TGF-β, recruiting MDSCs, and driving T-cell exhaustion through PD-1/PD-L1 and other inhibitory checkpoint axes. Even in patients who mount an initial vaccine-induced T-cell response (low magnitude, below the threshold), maintaining effector function within the tumor remains problematic. PDACs’ low tumor mutational burden (passenger mutations) limits the neoantigen repertoire for targeting. Another problem is HLA heterogeneity across patient populations, which limits the generalisability of any single peptide formulation. Logistical constraints such as tailoring personalized vaccines, which require high-cost genomic infrastructure; rapid turnaround time between surgery and adjuvant treatment, and carefully timed deployment of combined therapy to prevent lymphocytic exhaustion compound the biological ones. All these challenges limit scalability in routine clinical systems. The approximately 50% response rate, as reported by Rojas LA et al., further underscores the urgent need for predictive biomarkers to stratify patients and tumors for optimal outcomes.
Future Directions
The logical next step to combat fractional efficacy with the sole use of vaccines is to synergize the armamentarium with combination therapies. Pairing vaccines with PD-1 or CTLA-4 checkpoint inhibitors addresses the effector dysfunction that limits single-agent efficacy by relieving inhibitory signals at the point of T-cell engagement. Concurrent chemotherapy as in the atezolizumab-mFOLFIRINOX backbone used in the autogene cevumeran trial enhances immunogenic cell death and antigen release. This further amplifies the vaccine signal in an otherwise antigen-poor microenvironment. Biomarker-driven patient selection represents an equally critical frontier. Immune profiling to characterize pre-existing tumor-infiltrating lymphocyte populations and HLA haplotypes could further refine candidate selection. Longer-term preventive vaccine strategies for high-risk individuals-those with hereditary PDAC syndromes or high-grade precursor lesions, may shift the therapeutic window to a point at which the immune system still retains its full capacity to respond.
Conclusion
Pancreatic cancer vaccination is, at its core, an exercise in reprogramming biological probability converting an immunologically inert tissue into one that the adaptive immune system can recognize, infiltrate, and destroy. The clinical results accumulated between 2023 and 2026 are prima facie evidence for such an ambition in a chosen subset of patients, and how, when it is inscribed, it can durably subsist in immune memory, meaningfully lengthening patient survival. Yet this is still in nidifugous translation, with response rates of approximately 50%, modest cohort sizes, and the absence of phase 3 data, which demands necessary epistemic humility. What this new era of cancer vaccines ultimately offers is not merely a new drug class, but a new conceptual diction for oncology, one in which therapeutic potency is derived not from chemical cytotoxicity but from the precision and persistence of one’s own immune memory and resilience. As AI-optimized neoantigen platforms bloom and delivery technologies enable precise targeting of the site of action, the immunosuppressive barriers that have long bulwarked PDAC’s clinical intractability are eroded. The prospect of engineered, programmable immunity against one of medicine’s most formidable adversaries moves incrementally from aspiration toward the golden glory of treatable cancers.
References
- Rojas LA, Sethna Z, Soares KC et al. Personalized RNA neoantigen vaccines stimulate T cells in pancreatic cancer. Nature. 2023 Jun;618(7963):144-150. PMID: 37165196.
- Wang Y, Huang P, Li C et al.Therapeutic cancer vaccines in pancreatic cancer. Front. Immunol. 16:1674743.
- Zhang J, Xiao L, Zhang Y, Jin G, Zheng K. AI-powered vaccine breakthroughs: Targeting pancreatic cancer with neoantigens and combination therapies. Biochim Biophys Acta Rev Cancer. 2025 Nov;1880(6):189484. PMID: 41138796.

Author: A P
A medical trainee with an emerging focus on translational and clinical research, with interests spanning surgical sciences, neuroscience, pediatrics, and immunology. Her academic trajectory reflects an effort to integrate molecular innovation with clinically relevant disease models, particularly in complex and high-burden conditions. Her research experience includes work in genome engineering, specifically in prime editing, exploring its therapeutic potential in precision medicine. She has also contributed to oncological research examining cholangiocarcinoma with brain metastasis, focusing on its clinical course and diagnostic challenges. In parallel, her work investigating stoma formation as an independent risk factor for acute kidney injury reflects an interest in perioperative and systemic complications. Academically, she has contributed to case-based and review-driven scholarship, including a case reports and interdisciplinary review articles. Her evolving interests in neurology, pediatrics, and immunology reflect a broader inclination toward understanding disease across systems—from molecular mechanisms to clinical outcomes—while maintaining a disciplined, evidence-based approach to patient care. MBBS (MS4) ABVIMS Dr. RML HOSPITAL New Delhi



